




 19:30
19:30
 Hoàng Phong Nhã
Hoàng Phong Nhã



Tàu HMS Beagle là tàu thăm dò, khảo
sát của Hải quân Hoàng gia Anh. HMS Beagle dừng lại ở bờ biển Brazil,
Argentina dọc Đại Tây Dương (Atlantic). Tại đây, Darwin đã thu thập và
quan sát các loài thú phân phối từ vùng nhiệt đới Brazil đến cao nguyên
Pantagonia, phía nam Argentina và các hóa thạch của những loài thú đã
tuyệt chủng mà ông tìm được ở sườn núi, vực đất đá nhô ra cạnh biển.
Những dữ kiện này cùng các dữ kiện khác ông thu thập được sau đó ở phía
tây Nam Mỹ dọc bờ biển Chile và quần đảo Galapagos ở Thái Bình Dương đã
tạo cho ông suy nghĩ về sự biến dạng và liên hệ giữa các loài sinh vật
qua không gian và thời gian.
Trước khi đến Nam Mỹ, Darwin đã mang
theo sách để đọc và trong đó có sách của von Humbold. Ông rất ngưỡng mộ
nhà khoa học và nhà thám hiểm Alexander von Humbold về hành trình và
những nhận xét của Humbold về địa hình, môi trường và sinh vật ở Nam Mỹ.
Tuy vậy tư tưởng lãng mạn của Humbold về sự đồng nhất và hoà hợp của tự
nhiên trái với những gì ông thấy và trải nghiệm về đời sống và thiên
nhiên ở vùng này: sự đấu tranh sinh tồn không những xảy ra giữa các loài
sinh vật mà ngay cả trong xã hội con người qua sự đối xử dã man đối với
nô lệ ở đây mà ông có dịp chứng kiến tận mắt. Chế độ nô lệ là vấn đế mà
dòng họ bên ngoại của ông chống lại và muốn bãi bỏ. Tư tưởng về sự liên
hệ của các loài vật qua các quan sát, dữ kiện mà ông thu thập trong
chuyến đi và qua đạo lý của gia đình trong sự tin tưởng là nạn nô lệ mà
chủng tộc này đặt lên chủng tộc khác là hoàn toàn sai trái của ông đã
dần dần hình thành trong ông những hạt nhân của thuyết tiến hóa.
Qua các quan sát và dữ kiện thu thập của
ông ở Brazil, Argentina, Chile, quần đảo Galapagos ở Nam Mỹ, những câu
hỏi về sự liên hệ giữa các loài bắt đầu làm ông suy nghĩ. Sau khi ghé
Tahiti, New Zealand và ở Australia một thời gian để quan sát, thu thập
dữ kiện các loài sinh vật, đặc biệt là các loài đặc hữu ở lục địa này,
ông càng tin tưởng về sự liên hệ của các loài sinh vật và ý tưởng về
thuyết tiến hóa thật sự hình thành trong đầu ông. Sau chuyến đi năm năm
trên tàu Beagle, năm 1836, ông trở về Anh và trong thời gian ở Cambridge
ông viết lại nhật ký, các bài báo cáo khoa học và gửi cho Hội Động vật
(Zoological Society) các mẫu của các loài thú có vú, chim cho các nhà
khoa học chuyên môn nghiên cứu. Sau đó ông viết sách, biên tập các sách
của các nhà khoa học khác nghiên cứu về các dữ kiện của chuyến đi tàu
Beagle. Năm 1842, ông rời thành phố cùng với người vợ mà ông cưới ba năm
trước đó, đến một làng nhỏ gần London. Ở đây, trong những lúc rảnh rỗi,
ông suy nghĩ về thuyết tiến hóa và viết trong nhiều năm trước khi quyết
định xuất bản cuốn sách về thuyết tiến hóa vào năm 1859.
Trong một số quyển sách nổi tiếng của Darwin Các chuyến đi thám hiểm và tàu Beagle (Voyages of the adventure and Beagle, 1839), Cấu tạo và phân phối của các bãi san hô(The structure and distribution of coral reefs, 1842), Nguồn gốc các loài (On the origin of species, 1859), Nguồn gốc của con người, và sự chọn lọc trong mối liên hệ với giới tính(The descent of men, and selection in relation to sex, 1871), và Thể hiện xúc cảm ở con người và thú vật (The expression of emotions in man and animal, 1872), thì quyểnNguồn gốc các loài là có ảnh hưởng nhất qua sự trình bày cặn kẽ, và đầy đủ bằng các dữ kiện và quan sát để thuyết phục cho thuyết tiến hóa.
Sau đây là những điểm đáng chú ý trong cuốn sách Nguồn gốc các loài của Darwin:
Trong phần giới thiệu, Darwin nói rằng
những dữ kiện quan sát thu thập được trong chuyến đi trên tàu HMS Beagle
về sự phân phối và sự khác biệt giữa các loài sinh vật hiện tại và
trong quá khứ đã để lại ấn tượng lớn làm ông sbí ẩn nguồn gốc của các
loài sinh vật. Ông đã quan sát các loài chim Rhea (một loài chim không
bay giống đà điểu) ở Nam Mỹ, loài rùa, chim bói cá (mockingbird) và chim
sẻ (finch) ở quần đảo Galapagos trên các địa bàn sinh sống khác nhau
(các đảo chung quanh) của chúng. Thí dụ, loài chim bói cáMimus thenca về hình thể hơi khác nhau ở mỗi đảo nhưng cũng có sự liên hệ gần gủi với loài chim bói cá trên lục địa Nam Mỹ gần đó.
Sau đó, ông đề cập đến những nghiên cứu
của mình trong nhiều năm sau chuyến đi, thu thập thêm tư liệu để viết ra
bộ sách đi đến kết luận, giải thích về nguồn gốc các loài. Trong giai
đoạn gần xong, ông nhận đuợc thư của nhà sinh học Wallace đang ở
Indonesia, trong thư Wallace nói về các loài sinh vật mà Wallace quan
sát thu thập được và đi đến kết luận giống như ông về vấn đề nguồn gốc
các loài. Sau đó, Wallace gửi cho ông một bài viết về vấn đề trên và nhờ
ông gửi tới Hội Linnean để đăng trong tạp chí khoa học của hội (bài của
Wallace và vài phần trong bản thảo sách của Darwin được đăng trong tạp
chí). Bởi thế, thay vì còn vài năm nữa để xong bộ sách lớn, ông đã cho
xuất bản quyển sách nhỏ tóm gọn hơn về nguồn gốc các loài sinh vật.
Ông nói rõ là mặc dù vẫn còn nhiều vấn
đề chưa rõ, nhưng ông không còn nghi ngờ gì nữa, sau bao nhiêu năm
nghiên cứu khách quan, rằng quan điểm mà đa số các nhà sinh học tin vào
(dù trước đây chính ông cũng vậy) cho rằng các loài sinh vật được tạo
sinh ra độc lập với nhau là sai lầm, các loài sinh vật không phải bất
biến, và các loài gần nhau trong cùng giống (genus) có cùng tổ tiên hiện
còn hay đã tuyệt chủng. Cuối cùng ông cho rằng, sự chọn lọc tự nhiên
(natural selection) là cơ chế chính tạo ra sự khác biệt của các loài.
Chương I, ông bắt đầu bàn về sự khác
biệt các loài động vật (như chim bồ câu, bò…) hay thực vật sau nhiều thế
hệ qua gây giống bằng sự chọn lọc của con người để giải thích về sự
không bất biến của các sinh vật và trong những trường hợp nào thì sự
chọn lọc được thuận lợi thành công.
Chương II nói về sự biến đổi khác nhau
của các sinh vật trong tự nhiên ở nhiều nơi trên trái đất. Ông đưa ra
nhiều thí dụ về khó khăn trong việc xếp loại các loài sinh vật hơi khác
nhau, như các loài chim ở quần đảo Galapagos có sự biến dạng về ngoại
hình thì chúng là một loài hoàn toàn khác (species) hay chỉ là một biến
chủng khác (varieties) và những sự nhầm lẫn cũng tương tự như khi lai
giống mà không biết chúng cùng loại lúc khởi đầu. Darwin không tin rằng
sự khác biệt là do ảnh hưởng lâu dài của tình trạng vật lý khác nhau ở
mỗi vùng. Những vùng địa lý có khí hậu tương tự và cùng vĩ độ như Nam
Mỹ, Úc, Nam Phi lại có các loài động, thực vật hoàn toàn khác nhau.
Ngược lại, có một số loài ở một vùng lục địa này lại giống, tương tự hay
có liên hệ với các loài ở lục địa khác mặc dù khí hậu ở hai nơi khác
nhau. Thật ra, những trở ngại tự nhiên như dãy núi, biển giữa đất liền
hay các đảo ngăn cách sự di chuyển của các sinh vật, gây ra sự biệt lập
trong thời gian dài là yếu tố quan trọng trong sự khác nhau giữa các
loài ở mỗi vùng. Những giống (genera) lớn có nhiều loài (species) phát
triển nhất là những giống có nhiều biến dạng nhất không tùy thuộc vào
địa lý. Những giống lớn lại càng trở nên lớn thêm qua sự biến đổi khác
nhau của rất nhiều loài khác nhau trong giống và vì thế thiết lập thêm
các loài mới.
Chương III nói về sự đấu tranh sinh tồn
(struggle for existence). Sự chọn lọc tự nhiên được thể hiện trong sự
đấu tranh sinh tồn của các sinh vật. Ngoài môi trường tự nhiên, các sinh
vật phải thích ứng và cạnh tranh với các sinh vật khác. Sự xuất hiện
của các loài mới từ một loài không thể chỉ được giải thích qua sự khác
biệt giữa các cá thể mà chính là do hệ quả của sự đấu tranh sinh tồn. Do
sự đấu tranh sinh tồn này mà qua bất cứ sự khác biệt nào dù nhỏ giữa
các cá thể, thì cá thể nào được lợi trong một hoàn cảnh sẽ sống sót và
truyền sự dị biệt thuận lợi này đến con cháu và nhờ thế chúng sẽ có
nhiều cơ hội sinh tồn hơn. Sự đấu tranh sinh tồn được hiểu theo nghĩa
rộng, cạnh tranh giữa các cá thể cùng một loài và giữa các loài không
những để sống mà còn sinh sản nối truyền. Thí dụ, cây tầm gửi
(mistletoe), một loài cây ký sinh sống bám trên các cây khác, các cây
tầm gửi bám trên cùng cành cây cạnh tranh lẫn nhau, nhưng cũng có thể
nói vì hạt cây trong quả của chúng được chim truyền đi để mọc chỗ khác
nên chúng cũng cạnh tranh với những cây khác có quả mà chim đến ăn. Tất
cả các sinh vật đều liên đới với nhau trong một mạng lưới đời sống và
tiến hóa dẫn đến sự đa dạng sinh học.
Các chương tiếp sau trong sách, Darwin
giải thích và cố gắng trả lời các câu hỏi khó khăn mà thuyết tiến hóa
gặp phải như tại sao có nhiều loài được coi là có liên hệ gần nhưng lại
không có các loài chuyển tiếp về đặc tính và hình thể ở giữa chúng
(Chương VI). Ông cũng đề cập đến bản năng trực giác (chương VII) của các
loài (thí dụ như chim cu tìm tổ chim khác để đẻ trứng). Ông không cho
rằng bản năng trực giác được truyền từ thói quen và đưa ra lý giải
thuyết tiến hóa về bản năng trực giác (instinct) của các loài vật như đà
điểu, kiến và ong: bản năng là di truyền thong qua sự chọn lọc tự nhiên
được thể hiện trên các bằng chứng cho thấy các loài có liên hệ gần với
nhau mặc dù sống ở các vùng khác và môi trường khác đều có bản năng trực
giác giống nhau.
Ông đưa ra các kiến thức ông biết được
qua những kinh nghiệm về lai giống (hybridism) cây và các loài vật để
tạo ra các loài hợp chủng (Chương VIII). Hai loài khác nhau nếu được lai
giống thường cho ra các con không sinh sản được và hai loài gần nhau và
có nhiều điểm giống nhau khi được lai giống thường có các con mạnh khỏe
và sinh sản được. Các dữ kiện này không trái với thuyết tiến hóa qua
chọn lọc tự nhiên (trong thời điểm này, kiến thức di truyền chưa được
hiểu rõ tường tận, phải nhờ đến công trình của Mendel ở đầu thế kỷ 20 và
sự khám phá sau này về hệ di truyền DNA trong nhiễm sắc thể tế bào
người ta giải thích rõ và chứng minh thuyết tiến hóa là có cơ sở).
Ông dùng các bằng chứng qua địa chất
học, như các hóa thạch của các loài động và thực vật, thông qua tuổi rất
xưa của các tầng địa chất chứa hóa thạch để hỗ trợ cho thuyết tiến hóa
và đưa ra các lời giải tại sao chưa hay không có trong hóa thạch các
loài chuyển tiếp giữa các động thực vật liên hệ với nhay (Chương IX và
Chương X).
Cuối cùng, trong chương XI, ông trình
bày những vấn đề trọng tâm mà ông đã suy nghĩ nhiều năm để đưa ra thuyết
tiến hóa là sự phân phối các loài qua các vùng địa lý khác nhau như ở
các đảo và lục địa (Nam Mỹ, Úc) mà ông đã có dịp quan sát và khảo cứu
trong chuyến đi trên tàu HMS Beagle. Sự phân phối các loài không thể
giải thích được bằng sự khác nhau của môi trường, điều kiện vật lý. Thí
dụ, cùng một vỹ độ và môi trường nhưng những loài sinh vật ở biển dọc
phía Tây vùng Trung và Nam Mỹ lại hoàn toàn khác với các loài ở phía
Đông. Ít có loài cá, cua hay sò nào có điểm chung mặc dù chúng chỉ ngăn
cách qua một dãy đất nhỏ, eo đất Panama. Ông nhấn mạnh về sự quan trọng
của các rào cản cách ly, các phương thức phát tán trong sự sinh ra các
loài từ một loài, vì thế tất cả đều bắt nguồn từ một trung tâm kiến tạo.
Ông cho rằng, trước khi cách ly các đảo gần lục địa và ngay cả các lục
địa có thể đã nối liền nhau trong quá khứ và vì thế các sinh vật đã phát
tán đến những nơi này và sau khi cách ly, chúng đã tiến hóa độc lập với
nhau. Điển hình là ở quần đảo Galapagos trên Thái Bình Dương gần lục
địa Nam Mỹ, các loài chim sẻ (finch) ở các đảo đều có liên hệ với loài
trên đất liền nhưng ở mỗi đảo, chúng đều khác nhau ở điểm nổi bật nhất
là độ lớn của mỏ, từ loài có mỏ rất lớn rồi dần dần chuyển qua loài có
mỏ nhỏ hơn đến loài có mỏ nhỏ nhất mà qua sự phân loại về hình thái và
phong cách sinh hoạt của nhà động vật John Gould (nhà sinh học cùng thời
với Darwin) thì chúng thuộc các loài (species) khác nhau nhưng cùng một
giống chim sẻ. Ngoài ra, về thực vật, ông cho rằng một số các hạt cây
có thể chịu đựng được nước biển, và khi trôi dạt hay được chim mang từ
nơi này sang nơi khác sẽ sinh sôi nảy nở tiến hóa qua nhiều thế hệ trở
thành một loài khác thông qua sự cách ly ở một môi trường khác với nơi
xuất phát. Ông cũng cho thấy các loài thảo mộc ở vùng núi lạnh
Scandiavia, Scotland (Tô Cách Lan), Pyrénées, Siberia, Labrador (Canada)
và Mỹ giống và có liên hệ với nhau vì trong thời băng hà, tất cả vùng
trên đều phủ băng và các loài phát tán trên cả vùng trước khi thời kỳ
băng hà chấm dứt.
Chương XII tiếp tục nói về sự phân bố
địa lý của các loài, ông bàn về các loài vật sống ở nước ngọt như ở hồ,
sông với các hàng rào ngăn cản nhiều hơn so với các loài trên cạn như
đất, đồi, núi, đại dương. Những quan sát ông thu thập được ở Brazil, Nam
Mỹ đã đặt ra cho ông một câu hỏi là tại sao ở mỗi lục địa trên các sông
hay hồ, mặc dù có đủ loại chủng loài với các loài cá rất khác biệt
nhưng lại có sự giống nhau giữa các loài côn trùng, ốc…. Và ở hai con
sông trên cùng một lục địa thường lại có cả các loài cá giống và rất
khác nhau. Ông cho rằng có thể là do có lúc trong quá khứ hai con sông
trên cùng lục địa đã gặp nhau hay trong những thời kỳ lụt lội đã trao
đổi nước và vì thế có sự liên hệ giữa các loài cá ở hai con sông. Ngay
cả ở hai lục địa khác nhau, có một ít loài liên hệ là do chúng cùng xuất
thân từ loài cá tổ tiên nước mặn đến từ đại dương thích ứng với môi
trường nước ngọt ở sông. Riêng các loài côn trùng, có thể là do chúng
thích ứng với việc di chuyển từ hồ này sang hồ khác, dòng nước này qua
dòng nước khác và vì thế phạm vi phát tán của chúng rất lớn. Và quan
trọng hơn, qua quan sát các rong nước do vịt mang đến và các trứng từ
chân vịt trong các hồ nuôi cá, Darwin đã cho rằng để cho là các loài vịt
trời, chim bắt cá đã mang trứng của các loài sò ốc dính trên chân của
chúng từ sông hồ này đến sông hồ khác.
Sau đó, ông trở lại vấn đề sinh vật trên
các đảo. So sánh trên các lục địa trên cùng một diện tích thì ở đảo số
lượng các loài bản địa khác nhau ít hơn số lượng trên lục địa (như New
Zealand với Australia hay Nam Phi) trừ những đảo có nhiều sinh vật do
con người mang đến. Nhưng ở đảo lại có rất nhiều các loài đặc hữu
(endemic) hơn trên lục địa, như số lượng loài chim đặc hữu ở quần đảo
Galapagos so với ở Nam Mỹ hay số lượng loài sò đặc hữu ở đảo Madeira so
với Phi châu, Âu châu. Ông cho rằng điều này chứng tỏ thuyết tiến hóa
của ông có cơ sở hơn là thuyết sáng tạo (independent creation). Ông cũng
giải thích các loài ếch, cóc thường không có trên các đảo là do trứng
của chúng không chịu được nước mặn và vì thế không phát tán được và nếu
thuyết sáng tạo đúng thì tại sao lại không sáng tạo các loài này trên
các đảo.
Về các loài có vú sống trên đất liền thì
ông chưa bao giờ thấy có ở các đảo xa lục địa hơn 300 dặm và ở nhiều
đảo gần lục địa cũng không có. Chỉ có các loài dơi (có vú) là có ở đa số
các đảo như New Zealand, Norfolk, Bonin, Mauritius, quần đảo Caroline
và Mariannes, Viti và các đảo này đều có loài dơi đặc hữu chỉ có ở nơi
đấy. Ông cũng đặt câu hỏi tại sao trên quần đảo Galapagos, ga6`n Nam Mỹ,
lại có nhiều loài sinh vật có liên hệ rất giống với các loài ở Nam Mỹ
mà lại hoàn toàn khác với các loài trên quần đảo Cape de Verde gần lục
địa Phi châu. Mặc dù hai quần đảo này giống nhau về khí hậu, đất (được
hình thành do núi lửa), độ cao rộng nhưng các loài ở Cape de Verde lại
có mối liên hệ với các loài ở Phi châu. Điều này không thể giải thích
được bằng thuyết sáng tạo nhưng lại phù hợp với thuyết tiến hóa của ông.
Chương XIII Darwin nói về sự khó khăn
trong sự phân loại (classification) các sinh vật qua hình thái, các bộ
phận cơ thể. Ta có thể tìm thấy dễ dàng các đặc tính chung giữa các loài
chim nhưng giữa các loài giáp xác (như tôm, cua,…) lại vô cùng khó
khăn. Ông bàn về môn hình thái học và phôi học qua đó các phôi (embryo)
của các động vật có sự giống nhau rất lớn về hình thái và sự phát triển.
Ông cho rằng sự khác biệt giữa các loài rất có thể là do sự thay đổi
nối tiếp sau này chứ không phải ở giai đoạn đầu và phân loại cũng nên
dựa vào các đặc tính của các phôi. Ông có kinh nghiệm nuôi và lai giống
chim bồ câu và thấy rằng khi còn nhỏ các loài rất giống nhau nhưng khi
càng lớn trưởng thành chúng lại càng khác nhau. Sự giống nhau của các
phôi cho ta thấy các sinh vật có cùng nguồn gốc nhưng về sau tách ra
thành các loài khác nhau.
Chương cuối cùng, Chương XIV, Darwin tóm
tắt và kết luận về thuyết tiến hóa của ông, những sự khó khăn mà thuyết
tiến hóa gặp phải (nếu như theo thuyết tiến hóa thì phải có các dạng
liên tục khác dần nhau trong tất cả các loài sinh vật nhưng tại sao ta
lại không thấy ở chung quanh ta) và những dữ kiện và lý lẽ ủng hộ thuyết
tiến hóa. Những lý do tại sao nhiều người tin vào thuyết sáng tạo, sự
bất biến của các loài mà chủ yếu theo ông là vì chúng ta chỉ nhìn vào
khoảng thời gian rất ngắn so với thời gian rất lâu dài qua tuổi địa chất
(geological age).
Sau đó, ông bàn về vấn đề thuyết tiến
hóa có thể được mở rộng đến đâu và hệ quả của sự chấp nhận thuyết tiến
hóa vào môn lịch sử tự nhiên (natural history). Ông cho rằng thuyết tiến
hóa sẽ ảnh hưởng đến nhiều ngành như địa chất, khám phá ra những sự
thay đổi di chuyển của đất, lục địa, đại dương cho thấy sự di cư của các
sinh vật và sự liên hệ các loài và đến các ngành tương lai như tâm lý
học, từ đó biết được nguồn gốc con người và lịch sử con người. Những
tiên đoán của ông một phần đã trở thành hiện thực mà chúng ta sẽ bàn chi
tiết thêm dưới đây.
Từ Darwin đến thuyết tiến hóa hiện đại
Đến đầu thế kỷ 20, thuyết tiến hóa của
Darwin đã lần lượt được củng cố, minh chứng và phát triển từ nhiều lĩnh
vực khoa học như cổ sinh vật học (paleontology) và các ngành mới như di
truyền, di truyền quần thể (population genetics). Sự khám phá ở
Solenhofen (Đức) vào năm 1861 (hai năm sau Darwin xuất bản Nguồn gốc các loài)
một hóa thạch “Archaeopteryx”, một loài sinh vật cuối thời Jurassic có
cánh, lông giống như chim nhưng lại có đặc tính của loài bò sát (có
răng, xương đuôi,…) đã làm Darwin và những người ủng hộ vững tin hơn về
thuyết tiến hóa. Một loài chuyển tiếp từ bò sát đến chim đã hiện diện
trước đây trên trái đất.
Tuy nhiên, vì giới hạn tri thức di
truyền trong thời điểm giữa và gần cuối thế kỷ 19 và cho là môi trường
ảnh hưởng trực tiếp đến đặc tính di truyền, nên Darwin đã đưa ra ý niệm
di truyền hòa trộn (blending) các tính trạng tập nhiễm (acquired
characteristics) qua thuyết mầm toàn diện (pangenesis), trong đó mỗi tế
bào trong cơ thể sinh ra các mầm (gemmules) tập trung vào tế bào giới
tính (tinh trùng và trứng) để di truyền đến thế hệ sau. Qua các hiểu
biết thêm về di truyền, Alfred Wallace và August Weismann đã bác bỏ ý
niệm Lamarck sai lầm này của Darwin và đưa ra thuyết di truyền dựa vào
thuyết tế bào mầm (germ plasm) của Weismann (1893). Các kiến thức mới về
hệ di truyền nằm trong nhiễm sắc thể (chromosome) hệ gen của tế bào qua
những kết quả nghiên cứu và khám phá của các nhà sinh học T. Boveri, W.
Sutton và T. Morgan giải thích được cơ chế di truyền các đặc tính qua
nhiễm sắc thể mà trước đây Gregor Mendel đã ghi nhận qua các thí nghiệm
về di truyền qua nhiều thế hệ.
Nhưng chính những công trình của các nhà
toán học Ronald Fisher, John Haldane và Sewall Wright về lĩnh vực di
truyền mà ngày nay ta gọi là di truyền quần thể đưa thuyết tiến hóa qua
sự chọn lọc tự nhiên đến một cơ sở khoa học có nền tảng vững và chắc
kiểm nghiệm được.
Bài nghiên cứu nổi tiếng của R. Fisher
có tựa đề “The Correlation Between Relatives on the Supposition of
Mendelian Inheritance” (Mối tương quan giữa các quan hệ họ hàng dựa trên
thuyết tiến hóa di truyền của Mendel) đăng trong tạp chí Philosophical Transactions of the Royal Society of Edinburgh năm 1918 được coi như bài viết quan trọng trong lĩnh vực di truyền và xác suất thống kê.
Bài viết này là kết quả của sự quan tâm
về di truyền của Fisher lúc còn ở Cambridge và sau này được sự hậu thuẫn
hỗ trợ của Leonard Darwin (con của Charles Darwin) sau khi Fisher tốt
nghiệp, rời Cambridge và lập gia đình, về quê sống trong thời gian Thế
chiến thứ Nhất (Fisher mắt kém từ lúc nhỏ nên không vào quân đội được).
Lúc ông được học bổng vào Cambridge năm 1909 cũng là thời gian không lâu
sau khi di truyền Mendelian được biết đến năm 1900 sau nhiều năm bị
lãng quên, không ai biết đến công trình nghiên cứu của tu sĩ người Áo
Gregor Mendel vào năm 1865. Lúc này ở Cambridge có hai trường phái khác
nhau về đặc tính của sự thay đổi di truyền trong quá trình tiến hóa. Nhà
sinh học W. Bateson thuộc trường phái Mendelian, tin rằng có sự thay
đổi lớn, bất liên tục của các đặc tính trong quá trình tiến hóa như kết
quả của Gregor Mendel cho thấy. Trường phái sinh trắc học (biometry) mà
nhà toán học xác suất K. Pearson đại diện thì lại ủng hộ ý của Charles
Darwin cho rằng chỉ có sự thay đổi nhỏ, liên tục dần dần trong quá trình
tiến hóa mà thôi.
Trong bài viết này, Fisher đã dùng toán
xác suất để chứng minh qua một mô hình di truyền là sự thay đổi liên tục
của các đặc tính có thể là hệ quả của di truyền Mendelian và vì thế
không có sự nghịch lý nào của kết quả di truyền Mendelian với ý tưởng
của Darwin về sự thay đổi nhỏ liên tục của các đặc tính trong quá trình
tiến hóa. Về phương diện toán học xác suất, Fisher lần đầu tiên đưa ra
khái niệm về phương sai (variance), tức bình phương của độ lệch chuẩn
(standard deviation) của các đại lượng độc lập ngẫu nhiên (independent
random variables) và từ đó phương pháp phân tích phương sai là cơ bản
cu/a toán thống kê và đã được xử dụng rộng rãi trong nhiều ngành toán áp
dụng.
Cũng trong thời gian này T. Morgan đã
phát triển thuyết của Bovery-Sutton về sự liên hệ của nhiễm sắc thể
(chromosome) với di truyền Mendelian. Qua các thí nghiệm trên ruồi, vai
trò của nhiễm sắc thể giới tính XY cũng có quyết định trong di truyền
các đặc tính. Mô hình gen trội (dominant), đồng trội (co-dominant), gen
lặn (recessive), kiểu hình (phenotype), kiểu gen (genotype), đồng hợp tử
(homozygote), dị hợp tử (heterozygote) và allele chứng minh rằng hệ di
truyền trong nhân tế bào là nguyên do của kết quả di truyền Mendelian và
đặt nền tảng cho thuyết di truyền cổ điển.
Mang định lượng vào di truyền, Fisher đã
đặt nền tảng cho di truyền quần thể và sinh thái học với các kết quả
toán học có thể kiểm chứng được. Ông cho thấy đột biến (mutation) gen
càng lớn thì xác suất độ khỏe (fitness) của sinh vật do đột biến mang
lại sẽ càng giảm và quần thể nào càng nhiều thì quần thể đó có nhiều sự
khác biệt do đột biến giữa các sinh vật và do đó càng có cơ hội sinh
tồn.
Cùng với các công trình của Fisher là
các công trình của Haldane và Wright, toán học về sự phân phối tần số
allele trong di truyền quần thể dựa vào khái niệm đột biến, sự chọn lọc
tự nhiên, dòng gen (gene flow) qua di trú hay nhập cư, độ lệch di truyền
(genetic drift) đã mang lại kết quả và giải thích thỏa đáng những quan
sát về hiện tượng trong sinh thái và cho thấy thuyết tiến hóa qua chọn
lọc tự nhiên là có cơ sở. Nhà toán học Mỹ S. Wright đã đưa ra quan niệm
F-statistic để xác định sự liên hệ và sự đa dạng (qua mức độ của dị hợp
tử) giữa các quần thể và trong quần thể. Ông đặc biệt để ý đến vai trò
của di trú liên quan đến độ lệch di truyền. Bộ sách đồ sộ của Wright Tiến hóa và di truyền của các quần thể
(Evolution and the Genetics of Populations) có ảnh hưởng lớn trong môn
di truyền quần thể mà các nhà khoa học cận đại về thuyết tiến hóa như T.
Dobzhansky, E. Mayr, và G. Simpson đã thiết lập và phát triển một cách
có hệ thống. Simpson dựa vào lý thuyết và kết quả của Wright về độ lệch
gen ngẫu nhiên (random genetic drift) để đưa ra thuyết tiến hóa với
nhiều tốc độ thay đổi khác nhau gọi là tiến hóa lượng tử (quantum
evolution). Đóng góp không kém quan trọng là quyển sách xuất bản năm
1932 tựa đềNhững nguyên nhân của tiến hóa (The Causes of
Evolution) của nhà khoa học Anh J. Haldane. Haldane là người đã khai
triển ra các phương pháp để ước lượng nhịp độ đột biến (mutation rate)
trong di truyền ở người.
Sau khi xuất bản cuốn Lý thuyết di truyền của sự chọn lọc tự nhiên
(The Genetical Theory of Natural Selection) vào năm 1930, tiếng tăm và
ảnh hưởng của Fisher rất rộng, ông viếng thăm các nước như Mỹ và Ấn Độ
để giảng dạy và tham gia nhiều buổi nói chuyện. Trong thời gian ông trở
lại Cambridge, ông nhận nhà nghiên cứu Cavalli-Sforza vào làm việc.
Cavalli-Sforza ngày nay là nhà khoa học nổi tiếng về di truyền ở người
và là người thiết lập ra đề án Đa dạng hệ gen con người (Human
Genome Diversity Project, HGDP). Nhà sinh học tiến hóa W. Hamilton,
người sáng lập ra thuyết tiến hóa chọn lọc huyết thống (kin selection),
cho biết cuốn Lý thuyết di truyền của sự chọn lọc tự nhiên đã làm ông say mê và có tác động lớn đến ông khi còn học ở Cambridge.
Các kết quả của di truyền quần thể cho
phép các nhà khoa học như J. Huxley, T. Dobzhansky, E. Mayr kết hợp với
các kết quả khám phá trong sinh học di truyền gen với thuyết tiến hóa
qua chọn lọc tự nhiên của Darwin để tạo thành một khung cơ chế Tiến hóa hiện đại tổng hợp
(Modern evolutionary synthesis) về thuyết tiến hóa qua chọn lọc tự
nhiên. Đây là phương thức mà hiện nay các nhà khoa học đều chấp nhận để
giải thích về thuyết tiến hóa của Darwin.
Julian Huxley, một nhà sinh học xuất
chúng và có cá tính nhân bản. Ông là cháu nội của T. Huxley (người bạn
luôn ủng hộ Charles Darwin), trong các thập niên 1940 và 1950 đã viết
sách nghiên cứu, tổng hợp các kết quả trong khoa học và nhiều bài trên
báo chí phổ biến về thuyết tiến hóa qua chọn lọc tự nhiên cho tất cả mọi
thành phần trong xã hội. Cuốn sách Evolution: The Modern Synthesis
(tiến hóa: Tổng hợp hiện đại) xuất bản năm 1942 đánh dấu thành quả kiến
thức khoa học về thuyết tiến hóa và có ảnh hưởng sâu đậm đến các nhà
khoa học trên thế giới. Được giải thưởng cao quý Darwin của Hội Hoàng
gia (Royal Society) và Darwin-Wallace của Hội Linnaean (Linnaen
Society), và được bầu là Tổng Giám đốc (Director-General) đầu tiên của
UNESCO 1946-1948, ông cũng là một trong những người thành lập tổ chức
Quỹ Hoang dã Thiên nhiên (World Wild Fund) năm 1961. Thuyết tiến hóa
hiện đại tổng hợp không những giải thích được các hiện tượng quan sát
định tính và định lượng trong các ngành sinh học qua quy trình tiến hóa
chọn lọc tự nhiên mà còn là nền tảng và công cụ cho các nhà khoa học
nghiên cứu tiên đoán và tìm ra những cái mới, chức năng mới trong các
lĩnh vực di truyền, hệ sinh thái, biến đổi gen, môi trường và ngay cả
các lĩnh vực khác như ngôn ngữ, thần kình, tâm lý, phát triển tổ chức xã
hội… có liên hệ đến đời sống.
Cùng với Huxley, T. Dobzhansky cũng có
những đóng góp không kém quan trọng trong sự phát triển di truyền quần
thể định lượng và tổng hợp với thuyết tiến hóa chọn lọc tự nhiên của
Darwin. Nhà di truyền học Mỹ (gốc Ukraine) Dobzhansky rất xác đáng khi
cho rằng Không có gì trong sinh học thấu hiểu được ngoại trừ dưới ánh sáng của tiến hóa(“Nothing
in Biology Makes Sense Except in the Light of Evolution”) trong bài
viết cùng tên. Ernst Mayr, nhà sinh học Đức, không dùng toán học như
trong di truyền quần thể nhưng cũng đóng vai trò tổng hợp di truyền
Mendelian, phát triển them thuyết tiến hóa của Darwin khi định nghĩa thế
nào là loài (species) và giải thích tại sao có sự xuất hiện của các
loài. Ông cho rằng sự biệt lập và tách biệt của một quần thể từ một quần
thể khác sẽ dẫn tới sự hình thành một loài mới khi loài này không còn
thể hòa giống sinh sản với loài kia nữa. Quần thể tách ra càng nhỏ thì
càng mau tiến hóa, dẫn tới một loài mới vì độ lệch gen sẽ nhanh hơn. Ông
đã giải thích thỏa đáng tại sao có sự phát tán, lan tỏa thích ứng
(adaptive radiation) của các loài mới từ các quần thể nhỏ biệt lập sau
khi di cư tách từ quần thể chính như trường hợp các loài chim sẻ của
Darwin ở quần đảo Galapagos. Ý niệm về phát triển chủng loài mới của
Mayr dựa vào quần thể nhỏ tách ra biệt lập khỏi quần thể chính
(allopatric speciation) là nền tảng cho Niles Eldredge và Stephen Jay
Gould sau này đưa ra thuyết tiến hóa theo cân bằng đột ngột (punctuated
equilibrium theory of evolution).
Khi Dobzhansky di cư sang Mỹ làm việc
với T. Morgan về di truyền ở ruồi thì không lâu sau ở Liên Xô, một nhà
sinh học người Ukraine, T. Lysenko bác bỏ thuyết tiến hóa của Darwin qua
chọn lọc tự nhiên và di truyền gen, thay vào đó Lysenko đưa ra một quan
niệm dựa vào thuyết của J. B. Lamarck. Chủ thuyết Lysenko đã bị Julian
Huxley phản bác là không dựa trên các phương pháp khoa học và không thể
kiểm nghiệm được. Với tư cách là nhà khoa học và đại diện cho tổ chức
quốc tế UNESCO, Huxley cho rằng ông phải có trách nhiệm phổ biến kiến
thức khoa học thông tin chính xác thực nghiệm chứ không cảm tính, cảm
nghiệm mang tính chất chính sách như của Lysenko cho mọi người. Ông miệt
mài không mỏi mệt viết sách, các bài viết trong tạp chí khoa học và báo
chí phổ thông phổ biến thuyết tiến hóa qua chọn lọc tự nhiên, phê bình
thuyết Lysenko chủ quan, mang ý thức hệ chính trị áp đặt vào khoa học tự
nhiên.
Năm 1954, một cuộc tranh luận khoa học
nổi tiếng ở Karachi (Pakistan) giữa Huxley và nhà sinh học Liên Xô N.
Nuzhdin (người ủng hộ Lysenko) nổ ra sau một hội thảo khoa học hằng năm
của Hội Phát triển Khoa học Pakistan (Pakistan Association for the
Advancement of Science). Trong hội thảo, Nuzhdin tấn công chỉ trích tất
cả những gì trong bài nói chuyện của Huxley về di truyền Mendelian.
Huxley thách thức Nuzhdin tranh luận ngày hôm sau về thuyết tiến hóa
Darwin – di truyền Mendelian và thuyết Lysenko-Lamarck với sự hiện diện
của tất cả mọi người trong hội và công chúng tham dự ở một hội trường
lớn.
Cuộc tranh luận này giữa Huxley-Nuzhdin ở
Karachi quan trọng và hào hứng không kém cuộc tranh luận gần 100 năm
trước ở Đại học Oxford năm 1860 giữa giám mục Wilberforce chống lại
thuyết tiến hóa của Darwin và người ủng hộ Darwin là nhà sinh học Thomas
Huxley. Kỳ diệu thay, Thomas Huxley ngày xưa cũng chính là ông nội của
Julian Huxley ngày hôm đó. Ngày tranh luận, hội trường đông nghẹt người,
tràn ra cả ngoài hành lang, các phóng viên phải đứng ngoài cửa. Cũng
như ông nội mình đã phục vụ cho khoa học và nhân loại trong buổi chiến
thắng vẻ vang ở Oxford, Julian Huxley đã hoàn thành nhiệm vụ không những
cho danh dự cá nhân của mình mà cho cả nền khoa học sinh học khắp nơi,
kể cả ở Liên Xô. Hôm sau, trên trang đầu của tờ báo The Star ở Karachi với tít lớn “Huxley versus Nuzdin: Huxley wins.”
Trong buổi tranh luận, Nuzhdin cho rằng
nguyên lý về gen chỉ là một tiền đề. Huxley trả lời “đó là một ý nghĩ
sai lầm thiếu hiểu biết. Gen và sự hiện diện của chúng đã được chấp nhận
là một tiền đề vì đã được suy ra từ những kết quả thí nghiệm, trước hết
ngay lúc đầu là những hệ số giả thiết, như Bateson đã làm, và sau này
thật sự là những đơn vị trên nhiễm sắc thể… các dữ kiện (của môn di
truyền ngày nay) đã được thu thập từ loài ruồi Drosophila và những loài
bò sát côn trùng (insects) khác, các loài có vỏ giáp (crustacean), từ
nhiều loài thực vật, men… từ các loài chim, các loài có vú, và cá và
ngay cả từ con người nữa. Giáo sư Nuzdin có thể không đồng ý với sự
kiện, ý niệm này hay kia nhưng ông hoàn toàn không có lý do để cho rằng
một lý thuyết toàn diện như thế là không có hiện hữu.”
Thuyết Lamarck ngày nay không được chấp
nhận trong khoa học vì không có cơ sở. Nhà di truyền và nghiên cứu nông
học xuất sắc N. Vavilov, người bạn và đồng nghiệp với W. Bateson, được
nhiều người kính trọng nhưng bị thất sủng vì ý tưởng của ông trái với
phương pháp và quan điểm của Lysenko. Sau khi Vavilov mất, năm 1968, ông
được khôi phục lại danh dự và Viện Kỹ nghệ Cây trồng Vavilov ở St.
Petersburg (Vavilov Institute of Plant Industry) được mang tên ông, nơi
trước đây ông làm việc và thu thập hạt giống từ khắp nơi trên thế giới,
ngày nay là một trong những viện nghiên cứu còn chứa nhiều hạt giống lớn
nhất.
Trong cuốn Nguồn gốc các loài, trong Chương XII, Darwin khẳng định
mối liên hệ giữa các loài trên các hải đảo và lục địa có thể được giải
thích qua thuyết của E. Forbes cho rằng trước kia các lục địa như Âu
châu và Phi châu đã trải dài ra xa nối liền với nhau và với các đảo (như
Azores, Madeira, Canary ở Đại Tây Dương), nhưng ông không dùng thuyết
của Forbes mà chú trọng vào thuyết tiến hóa qua sự di cư của các loài
đến các hải đảo. Sự liên hệ giữa sinh học và địa chất học được mở rộng
qua thuyết tiến hóa và thuyết trôi dạt lục địa (continental drift) vào
đầu thế kỷ 20 cho đến các thập niên 1950, 1960. Thuyết trôi dạt lục địa
do nhà khí hậu học người Đức, Alfred Wegener, đưa ra vào năm 1912.
Theo thuyết trôi dạt lục địa, cách đây
khoảng 180 triệu năm vào thời kỳ Jurassic, chỉ có một lục địa lớn gọi là
Pangea và một đại dương gọi là Panthalass trên trái đất và các sinh vật
sống trên cùng một lục địa Pangea này. Sau đó, Pangea bắt đầu nứt ra
chia làm hai lục địa Laurasia (gồm Bắc Mỹ, GreenLand, châu Âu, châu Á
ngày nay) và lục địa Gondwana (gồm Nam Mỹ, Phi châu, Australia,
Antarctica, Ấn Độ ngày nay). Cách đây khoảng 120 triệu năm, Gondwana bắt
đầu tách ra thành ba mảng lớn: Nam Mỹ-Phi châu, Ấn Độ và
Australia-Antarctica. Lúc này, loài thú có túi (marsupial) xuất hiện ở
Nam Mỹ (đi đến từ Bắc Mỹ thuộc Laurasia) và chúng di chuyển qua các dãi
đất (gọi là cầu đất, land bridge) lúc đó vẫn còn nối vùng cực Nam Mỹ với
lục địa Australia-Antractica trước khi Nam Mỹ, Australia và Antarctica
tách ra. Vì thế loài thú có túi đã lan ra chỉ ở ba nơi này mà thôi (cho
đến nay chúng vẫn còn tồn tại ở Australia và Nam Mỹ).
Cách đây khoảng 100 triệu năm, khi Nam
Mỹ và Phi châu nứt ra thì loài có vú xuất hiện ở Laurasia tràn xuống Nam
Mỹ, Phi châu và Ấn Độ (khi các lục địa này đụng với Laaurasia ở phía
bắc) cạnh tranh và khi chúng đến cực Nam Mỹ thì cầu nối ở đây với
Australia và Antarctica đã tách ra. Vì thế, hiện nay các loài thú có túi
còn nhiều nhất ở Australia (kangaroo, possum, koala, wombat, Tasmanian
devil) không bị tuyệt chủng hay còn rất ít như những nơi khác vì chúng
không bị cạnh tranh và hơn nữa nếu có loài có vú từ nơi khác đến thì
chúng vẫn trội hơn vì thích hợp hơn với khí hậu khô. Ở Nam Mỹ, vẫn còn
một ít các loài có túi (opossum) và ở Bắc Mỹ chỉ còn lại một loài
(Viginia opossum). Cách đây khoảng 60 triệu năm, Laurasia tách ra thành
Bắc Mỹ, Greenland, Eurasia, sau đó Bắc Mỹ nối với Nam Mỹ qua một eo đất
nhỏ. Cùng lúc đó mảnh Ấn Độ của lục địa Gondwana cũ trôi lên phía bắc
đụng với Eurasia tạo thành dãy núi Himalaya và loài có vú đến nơi này.
Lục địa Australia-Antarctica vẫn trôi biệt lập về phía đông nam và sau
đó Australia tách ra khỏi Antractica đi về phía bắc. trong khi
Antarctica dạt về phía Nam Cực giá lạnh làm cho mọi sinh vật trên lục
địa này đều tuyệt chủng hết.
Thuyết trôi dạt lục địa đã được kiểm
chứng qua nhiều hóa thạch động vật và thực vật và vì thế cho thấy sự
trôi dạt lục địa cùng với thuyết tiến hóa đã giải thích được sự phân
phối các loài liên hệ trên trái đất hiện nay. Đúng như Darwin đã tiên
đoán trong cuốn Nguồn gốc các loài, thuyết tiến hóa đã ảnh
hưởng đến địa chất học và minh chứng cho thuyết trôi dạt lục địa qua các
hóa thạch của các loài và sự phân phối của các loài sinh vật như loài
thú có túi ở Nam Mỹ và Australia.
Ngày nay, thuyết tiến hóa qua sự chọn
lọc tự nhiên đã có những tiến triển mới mở rộng phạm trù so với thuyết
tiến hóa của Darwin ban đầu. Sau đây là những vấn đề và những khám phá
mới trong lĩnh vực thuyết tiến hóa.
Tiến hóa theo cân bằng đột ngột (punctuated equilibrium evolution)
Như đã đề cập ở trên, phương thức tiến
hóa đến một loài mới qua sự chọn lọc tự nhiên có thể không phải là một
quá trình dần dần mà là thay đổi nhanh chóng đi đến một loài mới trong
một môi trường thay đổi đột ngột. Tốc độ tiến hóa trong trường hợp như
vậy sẽ rất nhanh gây ra sự tuyệt chủng của nhiều loài và sự xuất hiện
của các chủng loại mới thích hợp với môi trường đã thay đổi.
Năm 1972, trong bài viết “Punctuated
equilibria: an alternative to phyletic gradualism” (“Cân bằng đột ngột:
một phương cách khác so với sự tiến hóa dần dần của các loài”) hai nhà
cổ sinh học Niles Eldredge và Stephen Jay Gould đã thiết lập ra thuyết
tiến hóa qua cân bằng đột ngột dựa vào ý tưởng về sự sinh loài qua cách
biệt địa lý giữa hai quần thể (allopatric speciation) của E. Mayr để
giải thích những sự thay đổi đột ngột rất khác nhau của các sinh vật hóa
thạch trong các tầng địa chất. Các loài mới xuất hiện thay đổi rất
nhanh qua sự biệt lập, chúng không có cùng một không gian sinh tồn và
tiến hóa khác với loài tổ tiên và vì thế ta thấy các “khoảng cách”
(gaps) giữa các hóa thạch trong các tầng địa chất. Eldredge và Gould
dùng những kiến thức rất rộng trong nhiều ngành từ sinh học, toán học,
lịch sử đến triết học để giải thích sự tạo thành ý niệm sinh loài dần
dần mà nhiều nhà khoa học tin vào là sự thực và thuyết phục quan điểm
tiến hóa qua cân bằng đột ngột.
Trước đây, những “khoảng cách”, “gạch
nối bị mất” (missing links) là vấn đề gây khó khăn cho sự giải thích
thuyết tiến hóa và là những chứng cớ mà những người không tin hay hoài
nghi về thuyết tiến hóa của Darwin dùng để chỉ trích thuyết tiến hóa và
coi đó là bằng chứng của sự sáng tạo các loài một cách thình lình trong
phút chốc của “đấng sáng tạo”. Thuyết tiến hóa qua cân bằng đột ngột đã
đưa ra một giải thích thỏa đáng cho thuyết tiến hóa của Darwin về các
“khoảng trống” trong hóa thạch. Các nghiên cứu gần đây cho thấy sự thay
đổi khí hậu do con người gây ra qua khí nhà nóng đã đẩy sự tiến hóa đến
tốc độ nhanh chóng của một số sinh vật như sự phát sinh ra hai loài ếch
cây (treefrogs: Pseudacris crucifer và P. triseriata) rất khác biệt
trong sự trưởng thành và thích ứng trong hai môi trường khác nhau. Đây
là một trong nhiều bằng chứng cho thấy sự biến đổi đột ngột của các loài
đến một cân bằng mới là có thật và đã xảy ra.
Đứng trên quan điểm dựa vào khoảng cách
thời gian lâu dài thì tốc độ tiến hóa trung bình là chậm so với sự thay
đổi dần dần nhưng trong khoảng thời gian đó lại có những thời điểm mà sự
thay đổi xảy ranhanh chóng đột ngột. Thuyết tiến hóa qua cân bằng đột
ngột không trái với thuyết tiến hóa qua chọn lọc tự nhiên của Darwin mà
bổ sung cho thấy các loài đã được tạo ra như thế nào dưới sức ép của sự
chọn lọc tự nhiên.
Cũng không có gì lạ, S. Gould trong các
thập niên cuối thế kỷ 20, là một trong những nhà khoa học đã xông xáo
tích cực đứng ra tranh luận bảo vệ thuyết tiến hóa chọn lọc tự nhiên và
cân bằng đột ngột chống lại sự đả kích thuyết tiến hóa của các tổ chức,
cá nhân (chủ yếu là ở Mỹ) tin vào sự tạo thành muôn loài là do thần
quyền. Tuy vậy, nhiều ý tưởng mới khác của ông, như sự chọn lọc qua loài
(species selection) chọn lọc không qua thích ứng môi trường kiểu
spandrels (một loại đặc tính kiểu hình (phenotype) không qua đặc tính
kiểu gen (genotype)) đã không nhận được sự đồng thuận của một số các nhà
khoa học khác như John Maynard Smith, Richard Dawkins và Daniel
Dennett.
Một nghiên cứu gần đây của J. Whittal và S. Hodges về loài cây hoa bồ câu
(columbine Aquilegia) cho thấy chiều dài của ống cựa (spur) chứa mật có
khuynh hướng tiến hóa làm cho ống cựa càng dài thêm theo chiều hướng
của các loài côn trùng thụ phấn có lưỡi dài, và có các bằng chứng về sự
thay đổi đột ngột chiều dài ống cựa trong những biến cố tạo thành chủng
loài. Trước đây, vào năm 1862, để giải thích ống cựa chứa mật rất dài ở
hoa của loài thảo vật Angraecum sesquipedale, Darwin đã đặt ra
giả thuyết là có sự “chạy đua” đồng tiến hóa đã làm tăng chiều dài ống
cựa mật và lưỡi của côn trùng thụ phấn. Vì thế, ông tiên đoán là có sự
hiện hữu của một loài sâu bướm (moth) có lưỡi rất dài. Mặc dù sự khám phá loài sâu bướm Xanthopan morgani ssp. Praedicta
năm 1903 với lưỡi dài 22 cm đã kiểm chứng thuyết tiên đoán của Darwin,
nhưng mô hình “chạy đua” về sự tiến hóa chiều dài ống cựa chứa mật vẫn
còn nhiều tranh luận.
Kết quả nghiên cứu của Whittal và Hodges cho thấy loài cây hoa bồ câu Aquilegia
có các ống cựa chứa mật đã tiến hóa rất nhanh để thích hợp với những sự
thay đổi rất nhanh đã được định trước bởi sự thay đổi hình thái của các
côn trùng thụ phấn hoa. Những khám phá này cho thấy tiến hóa có thể
tiến triển theo con đường tiên đoán được và sự thay đổi nhiều nhất tập
trung vào lúc tạo ra loài mới.
Sự chọn lọc nhóm (group selection), chọn lọc huyết thống (kin selection)
Theo thuyết tiến hóa của Darwin và nhiều
nhà sinh vật học thì về cơ bản thuyết tiến hóa được thể hiện như sau.
Môi trường tự nhiên thường thay đổi. Tiến hóa qua sự chọn lọc tự nhiên
(natural selection) cho rằng sinh vật thích nghi với môi trường tự nhiên
sẽ tồn tại và phát triển và sinh vật không thích nghi sẽ bị đào thải và
tuyệt chủng. Sinh vật từ một tế bào đến nhiều tế bào (multicellular
organism) đều bị chi phối bởi luật tiến hóa qua sự chọn lọc tự nhiên và ở
phần vi mô thì tế bào và gen là tâm điểm của sự vận hành của quy luật
tiến hóa.
Tuy vậy, từ các thập niên cuối thế kỷ
20, một số các nhà sinh học đã nới rộng thuyết tiến hóa qua phạm trù
tiến hóa nhóm. Nhiều loài sinh vật thường sống chung trong một nhóm,
trong mỗi nhóm thì các thành phần đều có liên hệ gen gần gũi với nhau.
Ông Vero Wynne-Edwards trong thập niên 1960 cho rằng sự chọn lọc tự
nhiên tác động vào nhóm chứ không phải vào cá thể sinh vật và các cá thể
trong nhóm tự giới hạn sự tiêu thụ và sinh sản để cho cộng đồng sinh
tồn và phát triển. Cuốn sách của ông đã gây một làn sóng phản đối chống
lại ý tưởng của ông cho là tiến hóa tác động vào cộng đồng và vai trò
của cá thể là không đáng kể. S. Gould cũng là một trong nhiều nhà khoa
học hoài nghi về vai trò của nhóm trong quá trình tiến hóa của các loài.
Ý tưởng của Wynne-Edwards bị cho ra ngoài lề và quên rơi vào lãng trong
giới khoa học về thuyết tiến hóa, cá thể gen mới chính là tâm điểm của
quá trình tiến hóa. Điển hình cho quan điểm gen là tâm điểm, đơn vị của
tiến hóa là nhà sinh học R. Dawkins với cuốn sách nổi tiếng Gen ích kỷ(The
Selfish Gene, 1976). Dawkins, dùng các kết quả toán học của B.
Hamilton, cho rằng sự rộng lượng (altruism) của các cá thể trong nhóm
được hiểu và giải thích qua chọn lọc gen chứ không trên cơ sở chọn lọc
nhóm.
Tuy vậy, gần đây David Wilson cho rằng
gạt bỏ thuyết chọn lọc tự nhiên trên nhóm là một sai lầm lịch sử lớn.
Ông được ủng hộ bởi Edward Wilson, một nhà khoa học tự nhiên nổi tiếng
nghiên cứu về các loài động vật có tổ chức như kiến, ong. D. Wilson, O.
Wilson và một số các nhà sinh học khác cho rằng sự chọn lọc tự nhiên tác
động cùng lúc ở các tầng khác nhau: ở cả cá thể và nhóm.
Sự chọn lọc nhóm và sự chọn lọc liên hệ
huyết thống mà gần đây được nhiều người đề cập thật ra là tương đương
nhau, chỉ khác là đứng ở hai góc cạnh khác nhau của một ý tưởng. Sự chọn
lọc huyết thống cho rằng trong nhóm có sự hợp tác và nhiều cá thể đã tự
hy sinh, rộng lượng (altruistic) ngay cả có hại cho mình, để cả nhóm
được lợi – một số thể được hưởng lợi nhờ có hệ gen giống như gen của cá
thể hy sinh. Đây là thuyết của William Hamilton và được nhiều người gọi
là sự chọn lọc qua huyết thống (kin selection) để phân biệt với
ý tưởng tương tự là sự chọn lọc nhóm. Thuyết chọn lọc qua huyết thống
được thể hiện qua phương trình của Luật Hamilton (Hamilton Rule) như
sau: Sự rộng lượng sẽ xảy ra trong quần thể khi:
R x B > C
(R = Chỉ số liên hệ huyết thống, hay còn
gọi là chỉ số Sewal Wright, giữa cá thể cho và cá thể nhận, B = Lợi
điểm di truyền mà cá thể nhận thêm từ cá thể cho và C = Giá phải trả về
di truyền cho thế hệ sau mà cá thể cho phải chịu).
Thuyết chọn lọc huyết thống trở nên phổ thông và được nhiều người chấp nhận nhất là từ khi cuốn gen ích kỷ của Richard Dawkins xuất bản.
Một số các nhà sinh học như John Maynard
Smith, không đồng ý với thuyết chọn lọc nhóm (hay chọn lọc huyết thống)
và cho rằng trong nhóm chỉ cần có một cá thể ích kỷ, tăng trưởng không
hợp tác, hưởng lợi cho mình làm thiệt hại các cá thể rộng lượng là có
thể phá hoại sự thành công của cả nhóm. Vì thế, sự chọn lọc cá thể luôn
luôn thắng, lấn át sự chọn lọc nhóm.
D. Wilson và O. Wilson đồng ý là có sự
chọn lọc trong nhóm nhưng không nhất thiết là sự chọn lọc nhóm thua và
nhường cho sự chọn lọc cá thể trong nhóm mà tùy từng trường hợp. Họ tóm
tắt nguyên lý về sự rộng lượng trong nhóm qua ba câu ngắn gọn: “Ích kỷ
thắng rộng lượng trong nhóm. Nhóm rộng lượng luôn thắng nhóm ích kỷ. Tất
cả các điều khác chỉ là các ý kiến hay nhận xét”.
Trên tầng lớp khái niệm trừu tượng, thì
sự chọn lọc gen ở cá thể có thể không trái ngược với tiến hóa qua sự
chọn lọc tác động trên nhiều tầng lớp (multi-level selection). Samir
Akasha đã đặt ra một câu hỏi là sự chọn lọc nhóm có đòi hỏi nhóm phải tự
sinh sản như cá thể không hay là nhóm có các mức độ phát triển khác
nhau. Ông chia sự chọn lọc nhóm nhiều tầng ra hai loại gọi là MSL1 và
MSL2 (multi-level selection Type 1 and Type 2). MLS1 chú trọng vào vấn
đề sự phân chia nhóm có ảnh hưởng về số lượng, tần số của cá thể khác
nhau ra sao trong một quần thể. MLS2 chú trọng về số lượng, tần số của
chính các nhóm. Thí dụ cổ điển của MLS1 là chủ thuyết rộng lượng
(altruism): một quần thể gồm nhiều nhóm sẽ giúp nhóm rộng lượng sinh tồn
hơn là nhóm không rộng lượng và vì thể sẽ có nhiều cá thể rộng lượng
hơn. Thí dụ của MLS2 là các tế bào tạo thành nhóm trở thành một sinh vật
đa tế bào (multi-cellular organism). Các thành phần nhỏ kết cấu tổng
hợp thành các thành phần lớn, như từ các phân tử kết thành các tế bào
hay các “sinh vật” kết tạo vào một cộng đồng “xã hội”, để trở thành một
thực thể mới. Để các thực thể mới này bền vững thì khuynh hướng “ích kỷ”
của các thành phần phải bị đè nén ở tầng lớp nhóm. Nhóm nào mạnh là
nhóm có nhiều số lượng nhất.
Ông D. Wilson cho rằng sỡ dĩ những nhà
sinh vật học chỉ chú trọng đến vấn đề tiến hóa tác động trên cá thể gen
và không đồng ý với quan điểm tiến hóa qua sự chọn lọc nhóm thật ra là
bị ảnh hưởng từ bên ngoài lĩnh vực khoa học: “ý niệm cho rằng tư lợi
(self-interest) có thể là một nguyên lý lớn giải thích được nhiều sự
kiện trong mọi lĩnh vực dường như trở thành phổ quát vào khoảng giữa thế
kỷ 20, và những gì xảy ra trong lĩnh vực sinh học tiến hóa phản ánh một
phần của trào lưu đó. Hiện nay, trong bối cảnh của chủ nghĩa cá nhân và
tư lợi, chúng ta đang có một ý tưởng mới là loài người là một loài linh
trưởng (primate) tương đương với tổ ong hay một “sinh vật”. Vậy thì sẽ
có những hệ quả nào từ ý tưởng trên?. Đây không phải là khoa học tưởng
tượng: đây là sự đồng thuận ngày càng tăng lên trong cộng đồng các nhà
sinh vật học.”
Hiện nay, vấn đề về sự cạnh tranh hay
chiến tranh giữa các nhóm thúc đẩy sự phát triển của tính rộng lượng
trong các nhóm trong quá trình tiến hóa, được thảo luận rộng rãi. Chiến
tranh và rộng lượng là hai mặt của một đồng tiền. Một đề tài không kém
gay go là cá tính hành động (behaviour) của cá thể có phải nằm trong hệ
gen và di truyền đến các thế hệ sau hay không? Hay rộng hơn nữa là ý
niệm memes, các đơn vị ý tưởng (idea) trong văn hóa con người, của R. Dawkins cũng qua quá trình tiến hóa chọn lọc cácmemes trong xã hội loài người.
Các mô hình về tiến hóa nhóm (group
evolution) chỉ ra những hệ quả có thể giúp ta hiểu về sự phát triển của
xã hội con người. Sự chọn lọc nhóm là một trong những đề tài trong ngành
sinh học tiến hóa có ảnh hưởng trực tiếp đến việc số phận xã hội con
người thay đổi ra sao trong tương lai. Như ta đã biết trong sự chuyển
tiếp kết hợp thủy tổ của ty lạp thể (mitochondria) và của nhân (nucleus)
trong tế bào eukaryotic, tư lợi của cả hai được hợp nhất, nếu ta hiểu
thấu đáo về quá trình trên, ta cũng có thể hợp nhất tư lợi của các nhóm
trong xã hội con người.
Tiến hóa ở tầng lớp mạng gen
Ở tầng lớp gen, một kiểu gen (genotype)
thể hiện một loại đặc tính kiểu hình (phenotype) trên sinh vật. Trước
đây, người ta cũng cho rằng ngược lại một đặc tính kiểu hình (phenotype)
cũng chỉ được điều khiển bởi một kiểu gen (genotype). Trong bộ gen
(genome) của các sinh vật, mặc dù thuộc các loài khác nhau và hình thái
bên ngoài rất khác biệt, nhưng cũng có nhiều chuỗi DNA của một hay nhiều
gen giống nhau (và vì thế có cùng các protein tạo ra từ gen). Điều này
cho thấy qua hàng trăm triệu năm tiến hóa, một số gen cơ bản vẫn giữ
nguyên không biến đổi. Kỹ thuật xác định trình tự DNA (DNA sequencing)
và loại trừ gen (gene-knockout) cho thấy sự bao quát về bảo tồn của các
gen và chức năng của chúng ở các sinh vật. Thí dụ, các gen xác định sơ
đồ cơ thể của động vật hầu như hoàn toàn giống nhau về cả cơ cấu cấu
trúc và chức năng ở các loài thú, mà nếu nhìn bên ngoài thì chúng không
có điểm chung gì. Những khám phá như vậy làm vững hơn cho ý niệm trực
giác của mọi người là nếu trình tự gen và cách tổ chức của chúng được
bảo tồn thì các chức năng cũng sẽ được bảo tồn như vậy.
Nhưng gần đây đã có nhiều quan sát
nghiên cứu cho thấy điều này không còn đúng trong quá trình tiến hóa của
sinh vật. Có nghĩa là một gen có một vai trò trong một quá trình gồm
nhiều loại gen tạo ra một chức năng, nhưng một chức năng có thể có nhiều
hệ thống quá trình khác nhau chứ không phải chỉ có một hệ thống quá
trình. Mỗi hệ thống quá trình này gồm các gen khác nhau để tạo ra một
chức năng duy nhất giống nhau. Thí dụ, gen Bicoid ở ruồi Drosophila điều
khiển các gen khác thiết lập sơ đồ cơ thể đầu, cổ, bụng của ruồi trước
đây trong thập niên 1980 được coi là cơ bản và có trong tất cả các loài
côn trùng khác, nhưng những khám phá vào cuối thập niên 1990 cho thấy
gen Bicoid không có ở các loài khác như loài bọ bột mì (flour beetle) và
ong ký sinh (parasitic wasp) nhưng bù lại sự thiếu gen Bicoid lại có sự
thay đổi của các gen mà gen Bicoid điều khiển hoạt động để chúng phục
hoạt mà không cần gen Bicoid.
Có nhiều kiểu gen khác nhau qua các mạng
điều tiết gen (gene regulatory network) có chức năng cho ra cùng một
kiểu hình (phenotype). Mô hình phần mềm cho hệ thống gen cũng cho ra kết
quả như vậy. Như vậy, một kiểu hình hay một chức năng của một loài sinh
vật có thể có nhiều các dư thừa (redundancy) các gen tạo thành các hệ
thống gen mà thiết lập nên kiểu hình hay chức năng này của loài sinh vật
ấy. Thông thường chỉ một trong các hệ thống này được hoạt động ở một
loài nhưng ở một loài liên hệ có các gen tương tự lại không dùng hệ
thống đó để thể hiện kiểu hình hay chức năng mà lại dùng hệ thống khác
để thể hiện như các loài men làm phó mát là một thí dụ.
Dùng mô hình cho một kiểu hình nhưng với
nhiều kiểu gen khác nhau cho thấy khi kiểu hình được giữ bất biến không
đổi và đột biến (mutation) được cho phép xảy ra ở các kiểu gen thì sau
một thời gian được vận hành, mô hình sẽ cho ra một số lượng kiểu gen rất
lớn và trải ra rộng. Điều này cho thấy một loài sinh vật có một kiểu
hình nhưng có nhiều kiểu gen sẽ có lợi cho loài sinh vật đó trong quá
trình tiến hóa thích nghi với môi trường. Trong một thí nghiệm gần đây
của M. Isalan, trong đó 22 protein của vi khuẩnEscherichia Coli
được chọn và từ đó ông tìm ra hơn 600 dãy DNA khác nhau cho các protein
này. Các dãy DNA tổng hợp này sau đó được đưa vào bộ di truyền của vi
khuẩnEscherrichia Coli. Kết quả là khoảng 95% vi khuẩn có thêm
các gen DNA tổng hợp nhân tạo sống sót, điều này cho thấy mạng điều
khiển gen rất bền chịu được các DNA mới được mang nối thêm vào mạng di
truyền gen. Và một số các vi khuẩn này có thể cạnh tranh hữu hiệu hơn
các vi khuẩn bình thường khác.
Như vậy, sinh vật có thể có nhiều đột
biến mà vẫn bình thường, trái với ý niệm độ lệch di truyền đơn giản
trong di truyền quần thể. Không những thế, nó còn đặt lại ý niệm trong
ngành tiến hóa-phát triển (evo-devo) là các mầm sinh (embryo) của các
loài sinh vật giống nhau không phải vì chúng có cùng một gen kiểm soát
thiết lập sơ đồ mà vì có chung một đặc tính nào đó của các hệ thống mạng
gen điều khiển. Đây là một lĩnh vực mới mà thuyết tiến hóa đang được
phát triển và nghiên cứu chi tiết.
Tiến hóa dẫn đến thay đổi môi trường?
Sự cách ly của một quần thể ra các quần
thể biệt lập với sự thay đổi môi trường hay hệ sinh thái là nguyên nhân
và yếu tố quyết định dẫn đến sự tiến hóa từ một loài thành nhiều loài
khác khác nhau – một quá trình gọi là sự lan tỏa thích ứng (adaptive
radiation) của các loài.
Một câu hỏi quan trọng ngược lại mà ít
người tự hỏi là hệ quả của sự tiến hóa ngược lại có tác động thay đổi
môi trường hay không? Nghiên cứu gần đây cho thấy sự tiến hóa ra các
loài cá có gai ở lưng (stickleback) đã có ảnh hưởng rất lớn đến hệ sinh
thái nước ngọt trong các hồ ở Canada dọc bờ biển Thái Bình Dương. Loài
cá có gai ở lưng là loài cá đặc biệt không có vảy sống ở châu Âu, Bắc Mỹ
và Á châu. Chúng cũng có đặc điểm là con đực làm tổ ở đáy nước, ve vãn
con cái đến đẻ trứng trong tổ rồi thụ tinh trứng, sinh con và sau đó
chăm sóc các con.
Khi băng hà tan ở cuối thời kỳ Băng Hà,
cách đây hơn 10.000 năm, cá có ba gai ở lưng (stickleback) phát triển
trong các hồ nước ngọt được thành hình từ băng hà và tiến hóa thích hợp
với cách sinh hoạt và môi trường khác nhau. Ở một số hồ, có hai loài
hiện hữu qua quá trình tiến hóa sinh thái (ecological speciation), một
thích ứng với hệ sinh thái dưới đáy (benthic) gần ven hồ và loài kia
thích ứng với hệ sinh thái dưới sâu, xa bờ (limnetic). Ở một số hồ khác,
khi chỉ có một loài sống, thì chúng có khuynh hướng thuộc loại không
chuyên (generalist) vào một hệ sinh thái đặc biệt nào trong hồ mà sống ở
toàn các hệ sinh thái của hồ, mặc dù mức độ chuyên có thay đổi tùy ở
mỗi hồ. Qua các thí nghiệm thiết lập các hệ sinh thái trung tầm
(mesocosm) giống như ở hồ (thuộc hệ sinh thái vĩ tầm, macrocosm) trong
phòng thí nghiệm thì các kết quả cho thấy ngoài sự khác nhau của các
loài mồi (prey) như phytoplankton, zooplankton, ở mỗi hệ sinh thái khi
có sinh vật không xương (invertebrate) sinh sống và có sự hiện diện của
các loài cá có gai ở lưng (sống dưới đáy sâu gần ven hồ, xa bờ và phổ
biến khắp nơi) hay của cả hai loài (loài sống đáy sâu gần ven hồ và loài
xa bờ) thì môi trường ở mỗi hệ sinh thái cũng thay đổi khác nhau qua sự
hiện diện của các thành phần vật chất hữu cơ, và màu sắc cũng như cường
độ ánh sáng dưới nước đều khác biệt. Điều này là một bằng chứng rất cụ
thể chứng tỏ tiến hóa có ảnh hưởng và thay đổi hệ môi trường vô sinh
(abiotic) và sinh học (biotic). Môi trường thay đổi quá trình tiến hóa
thì ngược lại tiến hóa cũng thay đổi môi trường: cả hai hỗ tương nhau
kết cấu thành một hệ sinh học tiến hóa hoàn ngược (feedback).
Và ta cũng biết là bầu khí quyển chung
quanh trái đất có nhiều khí oxygen hơn so với lúc trái đất thành hình là
vì sự sản xuất oxygen kéo dài rất lâu cả tỉ năm do các vi sinh vật sơ
đẳng nhất xuất hiện trên trái đất như cynobacteria thuộc nhóm prokaryote
lấy năng lượng từ quang hợp thải ra oxygen và từ đó sự sống tiến hóa
đến các vi sinh vật sử dụng oxygen thuộc loài eukaryote. Đây cũng là một
bằng chứng cho thấy tiến hóa thay đổi môi trường trái đất một cách toàn
diện.
Tiến hóa trong lĩnh vực nhận thức, tâm lý và văn hóa
Thuyết tiến hóa của Darwin cho thấy, con
người và các loài linh trưởng cũng như các sinh vật khác có cùng nguồn
gốc. Từ sự có chung nguồn gốc lịch sử tiến hóa đó, ông tiến xa hơn nữa
và phỏng đoán cho rằng các sinh vật khác cũng có một số đặc tính, khả
năng nhận thức giống như con người. Đối với nhiều người thì điều này quả
là khó tin và cho rằng Darwin đã đi quá xa và cùng với các lý do khách
quan khác về thực nghiệm nên không được chú ý đến hay bỏ qua.
Tuy nhiên, trong nhiều năm qua đã có
những kết quả và câu hỏi liên quan đến mối lien hệ giữa khả năng nhận
thức của thú vật và con người như khả năng nhận thức ngôn ngữ, sáng tạo
âm nhạc, đồng cảm, huấn luyện… gây được sự chú ý và tranh luận của các
nhà khoa học về vấn đề mà trước kia Darwin đã nêu ra. Các nhà nghiên cứu
về tiến hóa trong nhận thức, ngôn ngữ và tâm lý như Daniel Dennett,
Steven Pinker cho rằng những khả năng này là do tiến hóa qua chọn lọc tự
nhiên. Con người có thể thể hiện những hình thức văn hóa vô hạn, thí dụ
như các thể loại nhạc từ baroque, hip hop, đàn đá, gamelan… cho thấy
những sự biến dạng như thế là liên tục, vô hạn trong khả năng cũng như
hình thức.Từ ý tưởng và quan điểm liên tục trên, sẽ không có các khoảng
cách nào trong sự phân phối khả năng nhận thức dọc theo tất cả các loài
sinh vật hay trong sự phân phối các hình thể văn hóa có thể có.
Con người có khả năng nhận ra các hình
dạng khuôn mặt khác nhau và các loài linh trưởng khác cũng có khả năng
này, cũng như khả năng “hòa giải” sau đụng chạm như ôm, hôn, vuốt ve …,
bắt chước, hoạch định tính toán, khôi hài, vui đùa. Các nhà khoa học
thần kinh khám phá các “tế bào thần kinh gương” (mirror neuron) trong óc
loài linh trưởng macaque và sau đó trong óc của chim sẻ, và thấy rằng
các tế bào thần kinh này xuất hiện ở tổ tiên của loài có vú và chim. Các
tế bào thần kinh gương phát tín hiệu khi con vật làm một hành động và
khi nó thấy hay nghe con vật khác làm hành động giống như vậy, tế bào
thần kinh gương được cho là có vai trò giúp con người có khả năng bắt
chước và đồng cảm.
Loài linh trưởng chimpanzee đã chết đuối
ở hồ quanh chuồng trong sở thú khi nhảy vào cứu đồng loại bị nạn. Nhiều
nhà sinh học, như F. de Waal, cho rằng sự nhận thức về đạo lý ở con
người là phát triển tiến hóa bắt nguồn từ thái độ, luật cư xử trong xã
hội các loài linh trưởng. Gen về đạo lý nằm ở hệ di truyền linh trưởng
mà con người là một thành phần.
Loài linh trưởng khỉ (tamarin monkey,
Saguinus oedipus) trong thí nghiệm gần đây đã phân biệt được văn phạm
tiền tố (prefix) và hậu tố (postfix) dùng cùng một âm tiết (syllable)
trong các từ có hai âm tiết trong tiếng Anh. Điều này cho thấy một phần
cơ chế văn phạm trong một số ngôn ngữ con người có chung nguồn gốc với
linh trưởng. Cơ chế này dựa vào nhận thức hay trí nhớ tiềm thức cơ bản
tiến hóa cho các chức năng khác không phải ngôn ngữ.
Tuy nhiên, quan điểm về sự liên tục
trong khả năng nhận thức không hoàn toàn nhận được sự đồng thuận. Nhà
sinh học M. Hauser cho rằng không thể có sự vô hạn liên tục các đặc tính
mà có khoảng cách vì sự giới hạn của các thông số trong sự phát triển
cũng như trong hệ gen các sinh vật có khả năng có rất nhiều dạng nhưng
chúng không được thể hiện hoặc không có trong nhiều loài. Những khả năng
nhận thức, sáng tạo ý tưởng trừu tượng, ký hiệu hóa… của con người là
những khả năng hiện nay chưa tìm thấy ở các loài sinh vật khác. Những
khả năng này có từ hệ thống tế bào thần kinh do sự biểu hiện của sự kết
hợp các gen mà hiện nay chúng ta chưa biết rõ chúng liên kết ra sao
trong bộ di truyền người là nguồn phát sinh đủ các “dạng văn hóa”
(cultural forms). Tuy nhiên, không phải các dạng văn hóa là liên tục và
vô tận mà chúng giới hạn trong “không gian văn hóa” (cultural space).
Thí dụ, ngôn ngữ có nhiều dạng và văn phạm khác nhau nhưng có những cấu
trúc ngôn ngữ văn phạm không thể nào có.
Kỹ thuật hiện nay có thể cho phép các
nhà nghiên cứu bắt đầu tìm hiểu về cơ chế tiến hóa về khả năng nhận thức
ở các loài vật. Thí dụ, các con chuột được dùng kỹ thuật di truyền để
ghép gen người FOXP2 (gen này có vai trò tạo ra tiếng nói), kết quả cho
thấy chúng học khả năng âm thanh (cụ thể là hành xử theo âm thanh) khác
các chuột có gen riêng của chúng. Đây là lĩnh vực nghiên cứu mới liên hệ
đến nhiều ngành khác nhau trong khoa học.
Thuyết đồng tiến hóa gen-văn hóa (gene-culture co-evolution theory)
Thuyết đồng tiến hóa (co-evolution) sinh
học, trong đó hai hay nhiều đặc tính sinh học của một loài hay giữa các
loài ảnh hưởng lẫn nhau và cùng tiến hóa (như loài chủ và loài ký sinh,
loài mồi và loài săn, cây hoa và chim hay bươm bướm, côn trùng) mà
Darwin có đề cập, được Paul Erhlich và Peter Raven khai triển thêm và
gọi là đồng tiến hóa trong bài nghiên cứu nổi tiếng vào năm 1964 khai
mào cho lĩnh vực nghiên cứu đồng tiến hóa, “Bướm và cây: một nghiên cứu
về đồng tiến hóa” (Butterflies and plants: a study in coevolution) (14).
Trong bài, Ehrlich và Raven cho thấy các cây khi tiến hóa sản xuất ra
các hợp chất hóa học chống lại được các loài bướm đến đẻ trứng, nếu
thành công sẽ phát tán đến nhiều nơi khác và thích ứng với môi trường
mới, tạo thành một loài mới. Nhưng sau đó bướm cũng sẽ tiến hóa thích
ứng với hợp chất này và cũng phát tán theo đến môi trường mới tạo thành
những loài mới. Đồng tiến hóa tạo thành chủng loài qua thoát và phát tán
thích ứng (escape and adaptive radiation co-evolution). Thuyết đồng
tiến hóa ngày nay đang có nhiều nghiên cứu và phát triển trong lĩnh vực
sinh thái.
Ngoài thuyết đồng tiến hóa sinh học của
Ehrlich và Raven, đáng chú ý gần đây là sự phát triển của thuyết tiến
hóa trong lĩnh vực văn hóa xã hội gọi là thuyết đồng tiến hóa gen-văn
hóa hay còn gọi là thuyết lưỡng di truyền (Dual Inheritance Theory).
Trọng tâm của thuyết đồng tiến hóa
gen-văn hóa là sự liên đới trong quá trình tiến hóa trong xã hội con
người giữa gen và văn hóa. Cũng như gen, văn hóa được truyền từ thế hệ
này sang thế hệ kế tiếp. Đơn vị văn hóa để truyền có thể là một ý tưởng,
ý niệm, niềm tin hay một biểu tượng (symbol) mang ý nghĩa, thông tin
trong xã hội, văn hóa của một quần thể hay như R. Dawkins gọi là “meme”,
một đơn vị thông tin (như phoneme, âm vị trong ngôn ngữ học) truyền
trong óc trong quá trình truyền văn hóa giữa các thế hệ.
Thuyết đồng tiến hóa gen-văn hóa được
đặt nền móng từ những công trình cơ bản của D. Campbell trong thập niên
1960 và sau đó phát triển trong các mô hình toán học đồng tiến hóa của
Marcus Feldman và Luigi Luca Cavalli-Sforza. Thuyết đồng tiến hóa được
biết đến nhiều sau khi R. Dawkins phổ thông hóa nó trong tác phẩm Gen ích kỷ (1976).
Cũng như di truyền quần thể qua gen, các
đơn vị văn hóa cũng chịu sự thay đổi ngẫu nhiên (random variation), độ
lệch văn hóa (cultural drift) tương tự như độ lệch di truyền ở quần thể
gen, các thiên vị trong văn hóa, sự chọn lọc nhóm văn hóa… đều có ảnh
hưởng đến sự truyền gen qua sinh học của quần thể. Trong thập niên 1980,
ba tác phẩm quan trọng về ý niệm và mô hình toán học trong di
truyền-văn hóa quần thể đã được xuất bản: C. Lumsden và E.O. Wilson với
công trình Gen, tinh thần và văn hóa (Genes, Mind and Culture, 1980), L. Cavalli-Sforza và M. Feldman với cuốn Nối truyền văn hóa và tiến hóa: Một tiếp cận định lượng
(Cultural Transmission and Evolution: A Quantitative Approach, 1981)
dựa nhiều vào toán học trong di truyền quần thể, R. Boyd và P. Richerson
với Văn hóa và quá trình tiến hóa (Culture and the
Evolutionary Process, 1985) cũng dùng các mô hình toán học và ảnh hưởng
các lực chọn lọc và thiên vị trong quần thể để giải thích các hiện tượng
tiến hóa trong quần thể con người.
Việc nghiên cứu và phát triển thuyết
đồng tiến hóa gen-văn hóa cho thấy ứng dụng và phạm trù của thuyết tiến
hóa đã được mở rộng qua các lĩnh vực khác ngoài sinh học.
Kết luận
Darwin đưa ra nền tảng lý thuyết tiến
hóa là sự thay đổi của các loài sinh vật qua thời gian từ trước đến nay,
bắt đầu từ sự thay đổi môi trường và sự đấu tranh sinh tồn dẫn đến sự
chọn lọc tự nhiên loài sinh vật thích hợp nhất. Thuyết tiến hóa đã trả
lời được câu hỏi quan trọng mà bao nhiêu người từ bao đời ở mọi xã hội
từ Đông đến Tây đã đặt ra là con người và các sinh vật từ đâu đến. Một
câu hỏi giản dị nhưng mang tầm triết học rộng lớn mà nhiều bộ óc của các
thế hệ đã cố tìm câu trả lời. Cũng như tư tưởng của Galileo cách đây
năm thế kỷ đã đảo lộn niềm tin trái đất là trung tâm của vũ trụ thì
thuyết tiến hóa của Darwin cũng đánh đổ quan điểm cho rằng con người là
một kiến tạo đặc biệt, trung tâm của muôn loài và trên các sinh vật khác
mà thật ra chỉ là một nhánh có liên hệ họ hàng gần xa với tất cả những
sinh vật khác trên trái đất kể cả các loài vi khuẩn đơn giản và cổ xưa
nhất nhưng vẫn còn hiện hữu hiện nay. Một khám phá có tính đột phá và
cách mạng trong lịch sử khoa học và tư tưởng con người.
Từ đó đến nay, trải qua 150 năm, thuyết
tiến hóa đã có nhiều bước tiến trong nhiều lĩnh vực khoa học. Thuyết
tiến hóa không những là nền tảng cơ bản trong sinh học, cổ sinh học,
sinh thái học để hiểu về sự sống, thế giới sinh vật trong quá khứ, hiện
tại, ở nhiều tầng lớp từ nhóm, cá thể đến gen trong nhân tế bào mà còn
phát triển đến những lĩnh vực khác như di truyền, di truyền quần thể,
sinh học phân tử, tâm lý, thần kinh, nhận thức học, văn hóa… Nói chung,
quá trình tiến hóa được vận hành và thể hiện trong bất cứ hoạt động tri
thức nào của con người. Thuyết tiến hóa vẫn còn đặt ra các câu hỏi khó
(siêu hình và không siêu hình) mà chưa được hay không thể lý giải được.
Nhận thức từ đâu và đến với quá trình thế nào. “Mục đích” của quá trình
tiến hóa là gì?, đi từ những sinh vật giản đơn đến các sinh vật càng
phức tạp với nhiều chức năng (có trái với luật thứ hai của nhiệt động
học không?), có thể tiên đoán được con người tiến hóa đến đâu không hay
thật ra không có một “mục đích” gì mà tất cả đều là ngẫu nhiên. Có các
đời sống nào khác trong vũ trụ ngoài trái đất không? Nếu có và nếu ngẫu
nhiên là luật phổ quát thì nơi đó có hệ sống đời sống thế nào?
Điều kỳ diệu của tiến hóa là một trong
kết quả hay sản phẩm các loài của quá trình tiến hóa là con người lại có
được sự nhật thức và khám phá ra chính quá trình tiến hóa này. Một sinh
vật hiểu được mình từ đâu đến và quá trình tạo ra mình cũng là một kỳ
diệu của tạo hóa.
Theo NGHIÊN CỨU LỊCH SỬ
 Posted in: Triết Học,Tư Tưởng
Posted in: Triết Học,Tư Tưởng

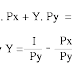



0 nhận xét:
Đăng nhận xét